前一页
回目录
回首页
个性的本原在何处?单凭一个人的脑,你至多能有把握地推测这个人是男性还是女性,但绝不可能确定这个男人或女人生前是否和善,是否有幽默感。正如我们在第一章中看到的,所有的脑都是按相同的基本模式构建起来的:有一些神经把感觉信息传进来,又有其他神经离开大脑,它们使肌肉收缩,并实施对运动的控制。我们也已经看到,脑是由神经元组成的,神经元在神经回路中活动,而这些回路一部分由遗传决定,但在相当大的程度上(至少是较复杂的脑)也受环境的影响。这种回路怎样转译为个体呢?在本章中我们将论述这个问题。
同卵双生者互为克隆。因为是由单个受精卵分裂而成,因此他们虽是两个人,却具有相同的基因。但他们完全一模一样吗?确实,用核磁共振(NMR)扫描对同卵双生的脑进行成象研究(参见第一章)的结果表明,在大体结构的层次上存在较大的相似性。如果向同卵双生者询问其爱好、态度和经历,答案常常也会有相当程度的相似,这也许并不奇怪。但是,在同样环境下抚育长大的兄弟姐妹在趣味和想法上的相重就没有那么显著。
同卵双生者也显示出不同的感知和思想的征象。这清楚地表明,尽管其遗传组成相同,但他们是具有其个人意识的个体。如果个性不能由基因来解释,那么它至少部分是由于大脑中存在某种其他因子,而这种因子甚至连起源于同卵的后代都不共有。
在前一章中我们已经看到,经验在塑造脑的微型回路中是怎样关键的一个因子。如果吃某种食物会让你联想起一件不愉快的事情,那你会厌恶那种特定的食物。举一个更简单的例子:只有听过莫扎特乐曲的人才会有可能说他们偏爱莫扎特。我们从未有过的经验不可能在形成我们的个性中起任何作用:如果某人具有学习多种语言的天生禀赋,却从未接触过不同的语言,那么其语言能力是不会显现出来的。
演变成一个独特的脑的过程在幼年至青少年期间可能最为剧烈,但是,即使在那以后,脑也并非一成不变。随着我们在成长的道路中连绵不断地遭遇经验和挫折,我们的性格继续与环境相适应。在这种方式下,经验显示出其持久的意义,因而它们就需要被记住。因此,个体的本质在很大程度上取决于其本身所记忆的东西。我们也许可以从记忆起步,来探索个性的物质基础。
至少在英语中,“记忆”(memory)是一个涵义甚广的术语,它所表示的各种过程可以迥然不同。试比较一下章鱼和人的记忆过程。章鱼的脑在无脊椎动物中是最大的,约与鱼脑相仿,由约1亿7千万个神经细胞组成。虽然这个数字看起来很大,但与人脑1千亿个神经细胞相比,则微不足道。尽管如此,章鱼被广泛地应用于学习和记忆的实验中,因为它有高度发达的眼睛和由众多触手组成的精巧的触觉系统。在实验中,一头章鱼能够清楚地显示对某些颜色的不同反应,并赋予每种颜色以不同的意义。例如,如果把一个颜色球和投食一头虾关联起来,让章鱼学会后,它就会轻易地抓住那个色球;而对另一个不同颜色的球,要是从未与任何奖惩刺激相关联过,它就不会作出任何反应。
这种类型的记忆(在一个色球和一头虾之间简单的关联),看起来好像与我们对海边炎热夏日的记忆,或记住如何骑自行车或记住法文“窗户”怎样讲,完全是两码事。“记忆”,这个总括性术语,所表达的大脑过程有许多不同的类型。在短期记忆和长期记忆之间的区别是最基本的,也是人们最熟悉的。当我们试图记住一串数字时动用的便是短期记忆。如果不分心,那没有什么问题,通常只需在脑子里一次又一次地重复这串数字就可以把它记住。令人惊诧的是,我们在这方面的能力相当有限:我们平均只能记住7位数。
关于短期记忆一个最明显的问题是,它和长期记忆有何相关?这类不那么费劲的记忆过程毋需重复或复述便能发生。那么,短期记忆和长期记忆是以平行的、完全独立的方式运转的吗?众所周知,有些病人对发生在不久之前的事毫无记忆,具有一种几乎是全面性的遗忘症状,但其短期记忆能力却与正常人不相上下。因此很清楚,这两个过程是可以分开的。但是,当某人丧失了短期记忆能力后还能有正常的长期记忆吗?
对短期记忆损害不易进行研究。长期记忆并非是一种一蹴而就的过程,正如我们在第二章中所看到的其他脑功能的情况那样,它能分成许多不同的侧面,对每一个不同侧面似乎都存在相应的短期记忆的形式。例如,一名对无意义的单词短期记忆不佳的幼儿,对不熟悉的玩具名称的长期记忆也差劲。短期和长期记忆似乎并非独立、平行地进行工作,而是以串行的方式实现的。首先是短期记忆开始运转:它是一种瞬间过程,高度不稳定,且易变。而为了导致更持久、更隐伏的长期记忆,需要集中注意力和重复。短期记忆的不断重复将最终使我们自然而然地记住某个特殊的电话号码,而毋需老想着它。
我们大家都知道,对于有意义的数字,如电话号码或楼房、保险箱的安全密码,短期记忆就会变得更好。在任何情况下,如果一项事情在你的记忆中保持约30分钟以上,那至少在几天之内你不会忘掉它。从脑震荡或电休克治疗(对严重抑郁症的一种根本性疗法)恢复过来的病人的特点是,记不起在这之前一小时前后发生了什么,但他们的长期记忆仍然有效。在这些情况下,有可能只是记忆过程的第一步,即短期记忆阶段受到了破坏。在事件的正常进程中,这种发生在早期的记忆中断,使在那个小时内所发生的事情不可能再在其头脑中较持久地记录下来。
短期记忆是为长期记忆服务的。但是我们说的长期记忆是什么意思呢?看来,这又是另一个与记忆有关的涵义广泛的基本范畴,其本身又能进一步分成两种不同的现象。在一生中我们要学习和记忆的东西真不少:如何开车;如何用法文讲“谢谢你”;当弗洛大婶上次来访时,我们做了些什么,等等。所有这些都是不同类型的记忆运转的实例。但在这三个例子中,与众不同的要算是如何开车了。对于一个事实的记忆(如法文“谢谢你”),或对于一个事件的记忆(如弗洛大婶近日的来访)需要我们作出明确的、有意识的努力。相反,开车,像许多技能与习惯一样,几乎是在自动引导下进行的。这类记忆因此称为隐性记忆,因为我们毋须主动、有意识地记住如何干某件事:我们只是坐上车驾驶而已。当你接近红灯时,你的脚会“自动地”去踩刹车。与这种过程相对照,对事实和事件的记忆被认为是显性记忆。
最著名的、研究得最充分的显性记忆完全丧失的病例之一是一个名字起首字母为H.M.的病人。他是一名青年男子,患有严重的癫痫,这种病的患者在发作时伴有意识的丧失。在H.M.这一病例中,病人癫痫发作得非常濒繁,致使他无法过正常人的生活。
1953年,H.M.27岁时,医生作手术切除了他大脑的一部分以控制癫痫的发作。尽管成功地治愈了癫痫,但是从此之后这种手术便不再进行,因为它造成了严重的后果:H.M.只能记得手术前约两年以前的事情。自手术之后,H.M.的意识总是停留在现在之中。
很难想象H.M.的思想状态。他不能辨认他在术后认识的朋友或邻居。虽然他能说出自己的生日,但讲不出自己的确切年龄,总是把自己估计得比实际上更年轻。夜间,他会问护士他在哪里,为什么他会在那里。他解释道:“每天都是孤零零的一天,不管我有过什么欢乐,或有过什么悲伤。”对H,M.来说不存在昨天。
这样的情况使H.M.只能完成现时的一些简单的动作。因此,只能让他干一成不变的活,如把香烟打火机固定在陈列板上。他无法描述他工作的地点、所干的活,也讲不清楚每天人们用车把他送来时所经过的路径。
但H.M.仍然能够记住7位数,这表明短期记忆是一个与长期记忆后续阶段相分离的过程。此外,虽然H.M.似乎失去了长期记忆的能力,他的大脑却保留了一种不同类型的记忆。H.M.实际上能相当好地执行某些运动性技能,如勾画五角星的轮廓。这种作业看来容易,其实并不然,由于是要求他对着镜子勾画轮廓,因此是一种很费劲的感觉运动协调的练习,这种练习越练越精,就像开车或骑自行车一样。H.M.每天在这方面都有进步,这就表明,处理另一种记忆――隐性记忆与处理事件的记忆并非在大脑同样的部位。饶有趣味的是,虽然H.M.的大脑对完成画星的作业越来越胜任愉快(隐性记忆),但他并不意识到是在记住学习画五角星这件事(一种显性记忆)。
虽然H.M.对发生在手术后,以及手术前两年间发生的事情毫无记忆,但对过去久远的事情的记忆仍然留在脑中不受影响,就像在琥珀中的苍蝇一样。这一点与我们眼下的讨论特别有关。这些记忆显然并不依赖于已被切除的脑区。可以肯定,没有一个脑区能够对事实和事件的整个记忆过程起全部作用。记忆必定是以某种方式通过一个脑区进行处理,而又在别处加以巩固的。在H.M.这一病例,损伤一定是发生在新记忆最初被处理的那个阶段,因此,所有已经巩固的记忆是保险的。正如我们在第二章中讨论感觉和运动控制时所看到的,不同的脑区负责处理一种功能的不同侧面。
H.M.被切除的脑区是其颞叶的中部,它位于大脑两侧,在颞颥附近,恰在耳朵的上方。这个区域也包括位于皮层下的称为海马(hippocampus,源于希腊语,因为有些人认为这个结构看起来像海马)的脑结构(参见图11)。在我看来,实际上最好是把海马看作一个位于皮层下的形状更像羊角的结构,围绕大脑的内实质卷曲起来。在H.M.的病例以后,又有许多临床和实验的证据表明,损伤这个脑区导致记忆贮存的损害。
 |
即使对于记忆的这个较特异的方面――记忆初始的巩固,也还有别的脑区似乎起重要的作用,那就是内侧丘脑,这个区域对于把传入的感觉信息转递至大脑皮层是至关紧要的(参见第二章)。正像听觉和视觉信息的处理各在丘脑不同的部分进行一样,丘脑中有一个特殊区域是与记忆有关的。
我们之所以知道内侧丘脑与记忆有关,是因为曾经发生过几起不幸的、怪诞的意外事件:击剑的剑具或台球杆捅人鼻孔,因而损毁了内侧丘脑。在这些病例中,意外事件的受害者表现出对事件的记忆不能。但是,与我们迄今已考察的遗忘症病例不同,他们的问题常常只是暂时的。尽管遗忘只是暂时的,但是对遗忘持续期间所发生的事情将出现永久性记忆不能,这可能正是内侧丘脑失去功能的时候。因此,就像海马一样,我们可以认为内侧丘脑对于记忆的巩固有着重要作用。
原始性遗忘是对一起事件发生在何时何地记忆的丧失。如果不存在空间或时间参照系,就不能对事件加以区分,也就不会有该个体对亲历已发生的事件的记忆。因为事件是独有的、个性的,而事实是普适的,与时间和空间参照系无关的,那么可以得出推论:原始性遗忘主要影响的是对事件的记忆而并非对事实的记忆。对事实和对事件的记忆似乎都依赖于海马和内侧颞叶的完整性,但只有对事件的记忆才受这第三个脑区――前额叶损伤的影响,我们在第一章中曾谈及这个脑区。
内侧丘脑与前额皮层有联系。有意思的是,内侧丘脑的损伤也能导致记忆的时空配置出现特殊类型的差错。记忆会不适当地突现出来,与当时的言语和思想毫无关系。前额皮层不仅可能影响事件所记忆的方式(如发生在某一时间和地点),还影响这些事件如何与可能在相近时间或地点所发生的相关事件的关联。
我们需要区别语义性记忆中的事实与经历性记忆中的事件,在后一种记忆中,事件是从一个特殊的时刻和地点取出来的。如果去年暑期的一晚你曾在丛林的幽处看到“红象――,那么一旦“红象”动了位置,你就会把它加以泛化――象可以是粉红的。在某个脑区,通过对时间和空间的参照,事实对那个人来说转化成了事件。要是损伤了那个脑区,所破坏的将不是记忆本身,而只是使事实与它所发生时的情景相脱离。特殊的事件会归化为普适的事实,在时间和空间上不再有特殊或独有的特征。
如果前额皮层对于事件记忆的这类时空配置是必需的,如果像我们在第一章看到的,前额皮层在进化的过程中已显示了惊人的分化生长,那么可以作出推论:对事件的这类记忆在人类特别突出,因为人类的前额皮层大得不成比例,而其他动物的前额皮层则要小得多。对于其他动物,也许对一个事件的记忆更具普适性,并不那么与独特的时间和空间坐标挂起钩来。一头猫可能不会记住,在某一特殊的春日,在喝了一碟牛奶后,在爬上一棵树前,在后花园抓了一只老鼠,虽然它可能会对抓鼠这类事有较模糊、一般性的回忆。有趣的是,在一种特殊的、非自然情况下,我们人类的记忆也似乎更像这类普适性记忆。
这些先驱性研究是由怀尔德・彭菲尔德(Wilder Penfield,一名外科医生)于50年代中叶在加拿大进行的。彭菲尔德在500名动神经外手术的病人身上进行了实验。使人惊奇的是,大脑本身并无疼痛的感受,因此有可能暴露清醒病人的大脑而不使他们感到疼痛。在病人的同意下,彭菲尔德进行了非做不可的手术,来研究记忆在大脑中的贮存。因为大脑的表面是暴露的,病人又是完全清醒的,他就能对皮层的不同部分进行电刺激,并同时把病人叙述的当时感受记录下来。
大部分时间病人所叙述的并非任何新的感受,这也许并不奇怪。但有时会发生相当有趣的现象:病人声称他们能记起很生动的场景。他们常常说,这些记忆就像梦,是更加泛化的感受,并无特定的时空参照点。也许在这种高度人为的情况下,电刺激正好局部刺激了内侧颞叶,而并没有刺激其他对记忆必不可少、但离得较远的脑区。在这些远隔的脑区中,特别是前额皮层,在正常情况下对事件的记忆是起作用的。我们在第一章和刚才都谈到,没有前额皮层,我们的记忆还是存在的,但是更模糊,不那么特异,这也许与彭菲尔德的病人那种梦似的记忆相像,或者甚至和正常的梦相像。如果不管什么原因,前额皮层作用的减小确实会引起一种梦似的精神状态,那么可以推论,前额皮层不明显的动物并没有我们所具有的精确的记忆。它们的记忆会是一种缺乏时空背景、脱离现实的事实:对事件的“经历性”记忆几乎已变成了对事实的“语义性”记忆。
我们在第一章中已谈到,前额皮层在工作记忆中似乎起重要作用。在这种记忆中,传入的信息和进行的行为受到某些内在的个体性想法、感知或规则的影响,这些在一生中积累起来的内在资源组成了一个个性化的精神世界。这些内在资源会提供某种砝码,以平衡迸发性的感觉信息洪流对大脑的冲击。人们常常把前额叶皮层的损伤与精神分裂症相比较;反之,人们又把精神分裂症部分归咎于前额皮层功能的失调(参见第一章)。精神分裂症的一种显著的共同特征是,过分地注意外部世界,而这外部世界常常似乎是过于精彩、过于闹哄哄了,却没有清醒地对内在资源作出合理的阐释,并依据经验加以调整。也许梦幻者、精神分裂症患者和人类以外的动物都有一种相似的意识,其特点是对以前的事件只有少许记忆,而以普适性的事实和即时直觉占据优势。如果确系如此,那么这样一种知觉状态可能会在菲尼亚斯・盖奇前额皮层严重损伤后的性格变化(参见第一章)中起作用。
我们已经看到,就事件和事实的显性记忆而言,临床病例(如H.M.)揭示,海马和内侧丘脑对于约两年的记忆存贮起作用。正如彭菲尔德的研究所揭示的,这些长期记忆,以某种方式“存贮”在颞叶中。同时,与海马和内侧丘脑都有联系的前额皮层把事实与适当的时空环境协调起来,从而保证把一个事件作为一次独特发生的事情记忆下来。
一个事实或事件是怎样在大脑中存贮起来的呢?我们知道,即使对过去事件的记忆有可能在丘脑和海马损伤后仍保留下来,它们也远非是不可磨灭的。把因内侧颞叶的切除而引起的记忆丧失,如H.M.的情况,与另一群病人的另一种记忆丧失进行比较很有启发。这另一群病人在记忆上的问题是因慢性酒精中毒而引起的。大量摄入酒精有许多危险,其中之一是引起科尔萨科夫综合征(Korsakoff’s syndrome),这是一种与食物中缺乏维生素B1相关的疾病。这种疾病的患者不仅有和H.M.相同的记忆损害,即对手术后发生的事情的遗忘(顺行性遗忘),而且也丧失了对入院前发生的事情的记忆,甚至在发病之前的事情也遗忘了(逆行性遗忘)。
顺行性和逆行性遗忘的区别在70年代的一项研究中显示得十分清楚。在辨认30年代和40年代名人的脸方面,事实证明患科尔萨科夫综合征的病人比H.M.更糟糕。从科尔萨科夫综合征病人来了解记忆过程的困难在于,很难把记忆的损害与其他类型的思维过程分离开来。酒精中毒者大脑的损伤非常广泛,除记忆之外还有许多其他过程也受到影响。与H.M.不同,科尔萨科夫综合征病人的其他不少脑区也有损伤,包括皮层的大块区域。
是否存在一个特定的脑区以最终安置记忆呢?在40年代时,心理学家卡尔・拉什利(Karl Lashley)曾试图回答这个问题。拉什利训练大鼠在迷宫中作记忆测试,然后切除皮层的不同部分,来看是否能鉴定记忆印迹贮存在何处。使他大为惊奇的是,在切除皮层的不同部分后,在一个特殊脑区和一种特殊记忆的保留之间并不存在一一对应的关系。结果倒是切去的皮层越多,大鼠完成记忆测试的情况越糟糕,与切去脑区的特异性并不相干。也许这并不奇怪,因为是整个皮层在记忆的存贮中起着重要作用。
与拉什利从大鼠得到的证据相一致,彭菲尔德报道的临床病例也揭示,记忆不是简单地被存贮起来,不是直接安置在脑中的,而是像彭菲尔德的研究所显示的,记忆的模式更像是一串朦胧的梦。那么立即产生的问题是,记忆本身并不像录象带上高度特异的记录,与计算机的记忆也大相径庭。另一个问题是,如果彭菲尔德在不同时候刺激同一个脑区,是否会引起不同的记忆?反之,刺激不同的脑区是否能产生相同的记忆?至今还没有人能明确无误地显示这些现象,并用大脑的功能活动来解释它们。但存在一种可能性,即虽然彭菲尔德每一回电刺激的都是同一部位,但所激活的是不同的神经元回路,而每一个回路可能参与一种特定的记忆。同样,当刺激另一个部位时,彭菲尔德有时倒可能是激活了以前曾激活过的回路,只是触发点不同而已,但是,不管是哪个触发点,一旦激活的是同一回路,记忆就会是相同的。
对彭菲尔德的发现可以作这样的解释:记忆是以某种方式与重叠的神经元回路相关联的。一个神经元可以是若干不同回路的成员,在不同的情况下一个回路与另一个回路的差异在于神经元的特异组合。每一回路都对记忆现象有作用,因此,并不存在单一的脑细胞或专门的细胞群起全部作用;毋宁说:记忆是分布性的。生化学家斯蒂芬・罗斯(Stephen Rose)在进行小鸡的实验时曾得出这个结论。他对小鸡进行训练,使之违背自然癖好不去啄食小珠。
简单说来,罗斯发现,鸡脑的不同部分所处理和记忆的是小珠的不同特征,例如是大小,而并非颜色。正像我们在第三章介绍视觉过程本身时所看到的那样,对视见物体的记忆是以平行方式安放的,并不存在单一的记忆区,而是分布在许多脑区。根据所记住的模态,以及在某一时空场景下被触发联想的不同,在整个皮层上将会有不同层次的回路募集起来参与活动。因此,不难理解拉什利怎么会得出这样的印象:在记忆过程中实际上所有皮层都以某种方式在一起活动。
记忆在皮层中最初是怎样巩固起来的?我们已经看到,所有类型的记忆一开始都进入短期记忆的时相,这一时相转瞬即逝,很容易解离;短期记忆至多持续半个小时。与此形成对照的是H.M.这一惹人注目的病例,他虽然能完全回忆起其一生早期所发生的各种事情,但他却对手术前两年间的事情毫无记忆。对于海马和内侧丘脑要巩固记忆来说,不是几分钟的问题,而应该是相当长的时间。
没有人真正确切地知道海马和内侧丘脑如何进行活动,与皮层结合起来在几年间把记忆存贮起来,使之最终不再依赖于这些皮层下结构的完整性。一种颇有吸引力的观点认为,记忆是由颇具任意性的单元组成的,这些单元在某一事实或事件记起时第一次汇聚在一起。海马和内侧丘脑的作用是保证使这些分离的、以前不相关联的单元关联起来,以某种方式结合成一种融合的记忆。正如我们在罗斯的实验中所看到的――那只是小珠的颜色相对于形状的简单情况,也有皮层的不同部分的参与。因此,需要有某种机制把这些不同的远隔的神经元群募集起来,形成一种工作网络。
可以想象,表示某种记忆的皮层工作网络的聚合取决于与海马与内侧丘脑间不断的对话。然而,当网络建立起来以后(可能要几年时间),皮层下结构就变得不那么重要了,以致一种已建立的记忆最终能脱离海马而不受影响,并完全独立于海马,就像发生在H.M.身上的那种情况。打一个比喻,这就好比是脚手架,在大厦正建造时,把脚手架拆了会影响施工,但一旦竣工了,脚手架便成了多余。
如果事件和事实的显性记忆依赖于最初在皮层和一些皮层下结构间的对话,那么也许同样的安排也能用于技能和习惯的存贮:隐性记忆。某些习惯,如记一串数字,或在适当的情景下做某类运动,是毋需思考的,因而内侧颞叶损伤而致遗忘症的病人能表现得不错。而罹基底神经节疾患(如帕金森病和亨廷顿舞蹈症)的病人(参见第二章),在事实和事件的显性记忆方面似乎没有问题,问题在于,他们不再能依习惯执行合适序列的活动:把一种序列一次又一次地加以显示,在正常情况下已经能形成隐性记忆,但他们却老是搞不清楚序列中的下一个是什么。
习惯的一个常见例子是,我们能在合适的时候产生合适的运动,但患舞蹈症的病人却不再能这样做。例如,这种病人的一个特有动作是踢腿,这在棒球投球时可能是恰当的,但在商场中显然不合适。另一方面,帕金森病患者不再能完成序列运动,序列越复杂(如起立或转身),困难就越明显。在这两种很不相同的基底神经节疾患中都出现隐性记忆系统的差错,表现在产生运动的习性的不同方面(分别是情景和序列)的错误。
基底神经节并非参与隐性记忆的唯一脑区。某些记忆作业包含有条件化反应,这很像我们在本章起始部分看到的章鱼的情况:一旦一种本来是中性的刺激(如一只球)与一种有意义的刺激(如一头虾)关联起来,它便会引起反应。现在认为,某些有肌肉的即时运动参与的条件化是由头后部的小脑控制的(参见第一、二章)。例如,在兔和人身上,都有可能建立起眨眼与一种本来是中性的刺激(如铃声)间的条件化反应,只要把后者与一种引起眨眼的自然刺激(如吹气)关联起来就可以了。
我们能看到,与事实和事件的显性记忆中使用的脑区相比,参与习惯和技能的脑结构是不同的。关键的差异并不在于这些结构的本体,而在于它们与皮层的关系。内侧丘脑和海马与皮层有很强的交互性联系,而与基底神经节和小脑的联系则并不那么强、那么主要。纹状体是基底神经节在舞蹈症和帕金森病中起关键作用的部位,它接收来自皮层的输入,但并不直接把信号送回去。同样,小脑虽与皮层间接地相联,但并无直接的联系。一种诱人的假设是,这些脑区不像那些参与显性记忆的脑区,在某种意义上它们能更自主地进行活动。可以预期,这种情况会发生在像隐性记忆那样毋需注意或有意识的努力便能进行的活动中,因为这些活动并不需要老是以皮层的活动为参照,而我们已经知道,皮层在有意识的注意中起关键作用。如我们在第二章所论及的,一旦一种运动变得能自动进行(或通过在基底神经节的内在触发信号,或通过经由小脑馈送的感觉输入),皮层便能腾出空来执行其他的功能,如显性记忆――对事实和事件的记忆。
我们已经看到,记忆能分为不同的过程,而每个过程是由脑区的不同组合来实现的。但是,也许最神秘的一点对所有这些记忆过程却是共同的。我们知道,有些人能记住90年前所发生的事情,而90年后这些人机体中的每一个分子都已经更新了许多次。如果媒介记忆的长期性变化是在脑中连续不断发生的,那么它们又是怎样保持下来的?不管是哪个脑区,神经元又是如何把经验所产生的或多或少持久的变化记录下来呢?
我们一直是在采用自上而下的策略来考察记忆。为了回答这最后一个问题,我们必须自下而上追溯,想象参与任何一种记忆过程的一个突触。为简单起见,让我们把记忆的最简单的形式看作是两个原先无关的单元间的一种关联。同样,为简单起见,让我们把这些单元的每一个都用两个单一的细胞来表示。
在记忆过程中,两个原先无关的神经元会同时活动,而这种相重合的活动最终会有某种持久的结果,其持续时间比每个细胞最初活动时间都要长得多。有远见的心理学家唐纳德・赫布(Don-ald Hebb)所提出的情况是最容易想象的。他设想,当一个输入细胞X特别活跃而使靶细胞Y兴奋时,在X和Y间的突触会增强。赫布说的增强指的是,与其他和Y接触的更沉静的输入相比,这一突触在化学信号传递方面变得更加有效。这个观点是在前一章中我们已经看到的在发育过程中所发生情况的翻版:活动最强烈的神经元(此处是X)形成最有效的联系。
第二种更近时的设想提出了联系增强的另一种方式,即参与联系增强的并非直接是靶细胞Y,而是经由第三个细胞Z。这第三个细胞在它向Y传送信号之前先影响X,因此,这种增强发生在突触前,而不是赫布模式中发生在突触后。如果Z和X同时活动使Z调制X的活动(参见第三章),那么就有更多的递质释放至最后的靶细胞Y。只有当X和Z同时活动,X才会随后向Y释放更多的递质(参见图12)。
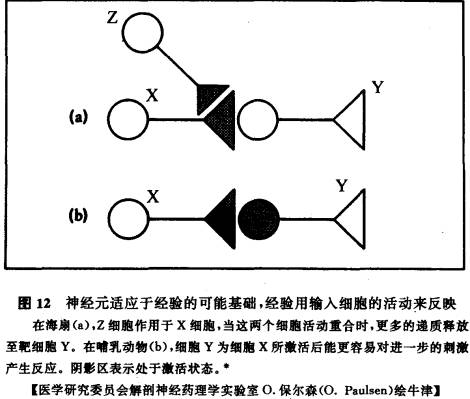 |
这种情况在海扇(海蜗牛)上已得到最成功的证明,海扇的优点是神经系统简单得多,甚至能对单个神经元加以鉴定、命名。在海扇较简单的神经系统中,把自上而下和自下而上的策略结合起来的问题并不存在,神经元回路的活动可以直接转译为可显示的行为。举例而言,一个神经元Z(可与上述的Z相比)对尾部的一种自然伤害性刺激有反应,它影响感觉神经元(X),后者对无害刺激有反应。这个感觉神经元又直接和用来缩鳃的运动神经元(Y)相联。
我们能够在海扇建立起条件化反应,使其在对感觉神经元给予一种中性刺激时缩回鳃,很像我们能在眨眼和铃声的中性刺激间建立条件化反应那样。当Z和X同时活动时,即当无害刺激和伤害刺激同时发生时,Z在X神经元诱发连锁性化学反应,导致钾通道关闭(参见第三章)。当这种荷正电离子的外流被阻遏时,跨细胞膜的电位就变得更偏正性,这正是使特殊的钙通道打开,并使钙进入细胞所必需的电压变化。当更多的钙因此进入细胞后,就有更多的递质释放出来(参见第三章)。感觉神经元X向运动神经元释放更多的递质,意味着运动神经元的活动变得更活跃,所引起的行为――缩鳃也就变得更强烈。神经元X,甚至在Z中止活动后,仍能维持在这种增强的状态。这样,条件化行为就建立起来了。
同样,在哺乳动物的脑,在许多参与记忆的脑区中的许多突触部位,都有可能存在某种高度活动的突触的增强。造成这类情况的一种关键机制称为长时程增效(LTP)。长时程增效是利用某种递质(谷氨酸)的一种靶受体(NMDA,即N-甲基-D-天冬氨酸)很挑剔的性质起作用的。与第三章中介绍的正常情况不同,这种受体只有在两个条件满足时才触发离子通道的打开。第一个条件和正常情况时相同,输入细胞必须也活动,从而使涉及的递质(在这种情况下是谷氨酸)能释放出来与其受体相结合。第二个条件是一种不寻常的要求,即该细胞必须已经具有比通常情况更偏正的电压。只有当这两个要求都满足时,那挑剔的受体才会让大量钙涌入靶细胞。
这两个要求只有当神经元活动重合时才能实现,这种重合活动可能通过以下两种方式发生。一种方式是,两个输入细胞同时活动,各自满足上述要求中的一个――一个输入细胞释放谷氨酸,而另一个细胞通过释放另一种递质而使电压降低。第二种满足这两个要求的方式则全和释放谷氨酸的那个细胞有关。最初,那挑剔的通道并不打开,因为虽然谷氨酸释放出来了,但细胞的电压处于正常水平。在正常情况下,那谷氨酸只是作用在其受体的不十分挑剔的亚型上。如果此谷氨酸的释放是持续性的,那么激活不十分挑剔的受体所产生的作用会使靶细胞的电压降低,从而满足第二个要求。那时,挑剔的谷氨酸受体能够打开钙离子通道使钙涌入。这样,持续性活动和输入细胞的相重合活动,在这两种情况下均能引起靶神经元长时程反应的变化。
输入神经元的这种持续性或重合性活动可以发生在记忆过程中。大量钙随后的流入又将在靶细胞内触发连锁化学反应,从而释放另一种化学物质,它越过突触,进入输入细胞,其作用是使后者释放更多的递质。靶细胞由此变得更活跃,这就是说突触增强了。当被增强突触的输入细胞又受刺激时(只要适度的即可),继后的反应将变得更大,这有点像海扇缩鳃的增强,称为增效。
这种现象可以解释短期记忆。但是,我们知道,短期记忆只持续不到一个小时。为了解释我们的似乎是永久性的记忆,必定要在细胞水平发生更持久性的变化。哺乳动物脑的长时程增效和海扇的增效一样,是一个必要的因素,但并非是充分的因素。如果递质释放的增多是持续性的,那么为了使短期内的反应变得更活跃或更强烈,递质必须把信号传送至突触另一侧的靶细胞,但这还不够。这种活动增强的持久结果实际上必须是发生在靶细胞内部的某种过程的变化。
很清楚,持久的变化不能简单地依赖于现存化学物质的大量释放。即使某些酶变得自发性活动起来,且它们所做的会增加突触的效率,但这些分子的寿命只有几分钟到几星期。虽然在记忆过程中细胞内发生的变化有许多还是谜,但有些事实正在显现出来。不管是海扇还是哺乳动物的长时程增效,我们都已看到,在这些现象中的共同事件是钙流入神经元。
这种钙的流入能起触发器的作用,在短至30分钟内,利用本身是短寿命的蛋白质来激活某些基因。这些基因的产物然后能激活别的基因,它们通过各种方式被表达,以此能很长时间地对神经元进行修饰。该神经元所激活基因的作用可以是增加递质的效率、受体的数目,甚或增加受体打开离子通道的效率。然而,还存在另一种甚至更根本的方式,使神经元可能为基因表达所改变。
我们在前一章谈到,经验所产生的效应并非那么显著地改变神经元本身,而是改变它们之间的联系。泛泛而言,经验越多,联系也越多。现已知道,若训练完成一种特定的作业,在一小时内,就会有某些重要的蛋白质活动起来。作为这些蛋白质的两个佳例,一是我们在前一章中谈到的细胞粘连分子,另一种是贴切地称为生长关联蛋白的GAP-43。细胞粘连分子看来对细胞识别和神经元间接触保持稳定起重要作用。随着细胞粘连分子在脑中产生,某些糖类将与之组合。如果给予适当的药物阻止糖的组合,就会产生遗忘症,这表明了细胞粘连分子在记忆中的重要性。
生长关联蛋白是在记忆中可能起作用的另一种蛋白质的实例,如其名称所示,它参与神经元的生长。生长锥(参见第四章)包含生长关联蛋白。我们已经知道,当神经元伸展其轴突时,生长关联蛋白的合成速率很高。在长时程增效过程中,生长关联蛋白显然被激活。因此,一个吸引人的推测是,在执行一种记忆作业时的神经元间接触的增强过程中,钙的流入,可能通过生长关联蛋白导致神经元接触的生长的增加,并可能借助于细胞粘连分子而使那些接触稳定下来。
通过这样一种方式就会产生新的突触连接,这是我们在第四章中谈到的在发育过程中的情况,也是脑反映环境变化最惹人注目的方式。在我们的一生中,与经验相适应的过程(即记忆)在我们脑中出现发育过程的回响,这并不奇怪。
神经元间联系的增加怎样导致记忆过程呢?这是一个很难回答的问题,因为它意味着我们要在哺乳动物脑中,在我们所讨论的自下而上的细胞层次和本章早先讨论的自上而下的功能研究间的鸿沟上架起桥梁来。我们需要知道,怎样把神经元尺度上的微观事件世界和记忆的宏观现象世界联系起来。虽然在海扇上,把某一神经元回路的活动转译为一种机械性行为(如缩鳃)是比较直截了当的,但在人脑,要把具有某种记忆的行为归结为一个特殊的神经元回路是不可能的。尽管如此,记忆过程的某些特征确实表明,神经元间的连接特性是十分重要的,虽然这些连接特性相当复杂,而且目前尚未得以鉴定。
为了提高记忆,一种众所周知的诀窍是把要记住的内容与某种本身会引起许多联想的东西挂起钩来。例如,把一个数字(“3”)与某种易想象的(“三只盲鼠”)和很熟悉的东西(一首古老的哄孩子的韵歌)关联起来,将使以后更容易回忆起这个数。另一种策略是,把譬如说一张购物单的各项东西想象是分布在房间的不同部位,如一块巧克力可以钉在门上,黄油可以放在台下,牛奶放在台上,茶放在洗水池里等等。还有一种改善记忆的方式是,使自己置身于或把自己想象为处于所记忆事件原来发生的相同场景中。为了记起你曾交谈过的救生员名字,你可以把自己想象是在暑期中海边沙滩上。做得更巧妙些,是去想象另一些东西,这些东西处于的场景就是所回忆事情发生的场景(如防晒液、毛巾、太阳镜等)。在所有这些情况下,我们或者是在巩固记忆时建立尽可能多的联想,或者在回忆过程中充分利用这些联想。
众所周知,大多数人记不起发生在约三岁时的事情。这种现象不能简单地用时间久远来解释,因为我们之后能记住的事情长达90年。此外,孩子们是能记住早年习得的习惯和技能的,因此问题只是在于显性记忆。另一方面,仅5个月的婴儿就能表现出显性记忆(这一点有争议),当两件东西一起显示时,他们更多地去看新的东西,而不是他们以前看到过的。不满一岁的孩子能依样玩他们前些天见到的别人玩过的游戏,即使他们只见到过一次。
看来,幼儿可能具有简单形式的显性记忆,这转而又意味着他们的海马和内侧丘脑一定是起作用的。就成熟程度而言,更成问题的倒是皮层。如果皮层神经元不能形成许多关联,那么幼儿的显性记忆就不会很强,实际情况也确实如此。这种把东西与通过经验积累起来的更丰富的信息源相关联的能力,在三岁以后才开始形成。这种能力,加上皮层中神经元间连接数目的增加,就使记忆成为可能。
虽然这些策略和例子可以不同,但基本的模式是相同的,即利用与已记住的东西间的关联。在神经元层次,这些关联肯定不是一种粗糙的单个细胞的一一配对。但是,在复杂性各异的神经元回路的庞大相互作用中,基本的变化单元将归结为我们所论述的连接特性的改变。我们知道,长期记忆伴有突触前终未数的增加;我们也知道,记忆包含着新的关联的建立。我们还不能在人脑中建立物质与现象间的因果关系,但是现下只需意识到两个运作层次间的相关就足够了。记忆是多侧面、多阶段的,不单是脑的功能,因为它充分利用了个体的内在资源,并以极独特的方式来阐述其周围的世界。正因为这样,以记忆来结束我们对脑的简短的论述可谓是恰到好处――它是我们精神的基石。
前一页
回目录
回首页