前一页
回目录
回首页
从一产生一切。
――赫拉克利特
分离,联合,变换,规定自身,显现与消失,固定与流动,伸展与收缩,是生命体的基本特征。
――歌德:《自然科学格言》
我们在自然界中发现巨大的组织结构,在这个组织结构中,下属的系统在连续的各层次上联合成更高的和更大的系统。化学的和胶体的结构整合成细胞结构和多种细胞,同种类型的细胞整合成组织,不同的组织整合成器官和器官系统,器官和器官系统整合成多细胞有机体,最后有机体又整合成超个体的生命单位。
这种组织结构的基础是物质的基本单位。按照物理学,物质由终极单位即各自带有负电荷和正电荷的电子和质子,以及不带电荷的粒子中子构成。它们构成化学元素的原子。由中子和质子组成的原子核好比微型行星系的太阳,一些像行星般运动的电子围绕着它旋转,每种元素则是由这些粒子特定的数量决定的。对这个系统的扰动,便使一种化学元素转变为另一种化学元素,而且可以导致原子的分裂,像原子弹爆炸那样毁灭性地释放出巨大的能量。
原子结合成为各种各样化合物的分子,部分属于无机界,部分属于有机界。尽管构成生物的化学元素与非生物的化学元素没有什么区别,但是,有机化合物是特殊的。由此引起了“无机”化学和“有机”化学的区分。在生物元素中,碳占有特殊的地位:似乎生命与碳的能力有密切关联,碳能形成极其多样、极其大量和极其复杂的分子,碳凭借这种能力而胜过所有其他化学元素。有理由说,作为碳化合物化学的有机化学与无机化学之间的区别,在于碳化合物的数量是所有其他化合物数量总和的许多倍。在有机分子中间,所谓大分子,例如,包括作为原生质最重要的构成物质和作为植物细胞壁纤维素的蛋白质,已经表现出复杂程度超过无机分子的特殊的结构规律。
无机和有机小分子量的分子,用精确的结构式表示,它们由原子价键的相互连接而成。正如人们所熟知的,一种元素的价键数是由能够结合在一起的氢原子数表示的。例如,氢具有一个氢价,H-H(氢分子);氧有两个氢价,
 (水);氮有三个氢价
(水);氮有三个氢价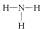 (氨);碳有四个氢价,
(氨);碳有四个氢价,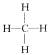 (甲烷)。
(甲烷)。
在这由精确结构式表示的分子的图像中,起初没有发生变化,当我们考察有机分子时,情况就不同了,正像我们在叶绿素、血红蛋白、维生素等大分子中看到的那样,从而结构式就变得复杂得多。
可是,我们一旦考察到高分子有机化合物,就出现新的结构原理。例如,如果我们考察植物纤维素,找们发现作为最小单位的二糖类,所谓纤维二糖;至少三百个这种“基本单位”由普通化学键连接成“主要化合价链””。然而,大量的这类化合价键,大约四十个到六十个,还要依靠第二类化合价或范德瓦耳斯力再连接成更大的结构――“胶态分子团”。蛋白质与多糖相对比,它们的区别在于,蛋白质不是由相同的,而是由不同的单位即不同的氨基酸构成的。在蛋白质分子的长链中,又出现确定的结构规律,发现这种结构规律是现代有机化学研究的主要问题。这些结构原则之一是,不同的氨基酸好像是按照周期性模式排列的。例如,在蚕丝的纤维蛋白中,每第二个链是甘氨酸,每第四个链是两氨酸,每第十六个链是酪氨酸。按另一个结构原则,许多蛋白质的分子量是35000分子量作为一单位的好几倍。
这里,我们可以指出三点普遍意义。第一,除了经典比学的化合价之外,还显示出有一种范围更广泛的力制约着物质的结合。这些力已在所谓非理想气体中表现为范德瓦耳斯力,范德瓦耳斯力由气体分子的相互吸引产生的,并造成偏离理想气体方程。它们作为晶格的力,在许多晶体形成的过程中起着构成作用,也表现为固体的内聚力。正如刚才提到过的,它们在大分子有机化合物结构中起作用,同样地,正如我们将要看到的,它们也在这些有机化合物形成胶态分子团单位中起作用。所有这些力原则上与化学的亲和力是相同的。可是,要指出的是,传统化学的基本的或主要的化合价只表示价键领域内相当小的一部分。在这个意义上,结构化学,气态、液态和固态理论,胶体化学,结晶学等等,融合成统一的领域。同时,无生命结构和有生命结构之间存在的鸿沟变小了。我们正洞悉着一个力的新领域,其中产生的构型和组织超越了传统化学仅仅考虑到分子内原子和原子团的构型。
第二,有序的类型本质上是变化的。尤其在大分子碳水化合物中,传统意义上的“分子”和“化合物”的概念变得不适用。事实已证明了这点。例如,在植物纤维素分子式(C6H10O5)n中,出现不确定的数n。植物纤维素的结构不能用严格的分子式表达,而只能用统计方法表达;大约三百个糖残基形成一个主要化合价链,大约四十个主要化合价链形成一个胶态分子团。
第三,胶态分子团又可以组合成更高级的结构。因此,例如植物纤维素胶态分子团在植物细胞壁中显示出有规则的排列,它们在连续的层次等级中最终逐渐形成微小的和肉眼可见的植物纤维。蛋白质中的层次等级尤为明显。氨基酸和蛋白质分子作为构成更高单位的部分,它们本身表现为细微的小纤维;小纤维又可以联合成微观纤维,而微观纤维又渐次联合成肉眼可见的宏观纤维,诸如神经和肌肉中的纤维。
这样,亚显微形态学领域(弗雷-维斯林[Frey-Wyssling」)形成了从物理-化学领域到生物学的转变。经典物理学和传统化学只知道两种有限的情况:一种是分子结构和三维晶格中原子或基团的严格排列;另一种是溶液中分子无规则运动的完全无序。但是,实际上物理结构的系列并不限于分子结构和晶体,而超出这些,只能应用溶液的随机分布定律和力学的克分子定律。相反地,这里出现了一系列更高级的、不间断地通向宏观领域的结构模式。在每一个新的结构层次上,自由度随之增加。分子结构是用精确的结构式确定的。像植物纤维素那样的大分子化合物,则只能用统计方法规定。在胶态分子团的排列中,诸如在小纤维中,其构成单位与典型的晶体相比,并不在三维空间上都是有序的,而只在二维或一维空间上是有序的。
超出大分子化合物的层次,就走向非生命界和生命界之间迷人的边缘地带。这是冠以病毒名称的病原因素的领域。小儿麻痹症、天花、麻疹、流行性感冒、狂犬病、口蹄疫等等,还有许多植物病,是由病毒引起的。病毒如此之微小,以致于大多数病毒不能用普通光学显微镜而只能用电子显微镜才能观察到。就简单病毒而言,它们具有巨大分子量的纯结晶蛋白质可以被分离出来。例如,烟草花叶病毒的分子量是40.7百万。另一方面,它们显示出来的属性似乎具有生命的最主要特征:它们通过分裂进行繁殖。例如,如果一株植物被注入几百分子的结晶烟草花叶病毒,那么,它的各个部分就会得病。病毒物质就会大量地增殖起来。
病毒,就其化学的和物理的性质而言,是可以用基本生物单位的概念加以概括的最典型的实体。这些实体被定义为能够复制自身或协变复制(covariant reduplication)的最小系统。病毒在许多方面可以与遗传单位或基因相比较,基因像一串珍珠那样排列在细胞核的染色体上,并位于染色体染色较深的片段上即染色粒上。然而,区别在于病毒像寄生虫那样是从外界导入的,而基因是细胞必不可少的组成部分。遗传学研究和染色体显微研究这两方面的工作得出了这样的结论:基因是由几十万分之一毫米长度的蛋白质大分子排列成的巨大分子链。因此,整个染色体可以看作是一种“非周期性晶体”(薛定谔「Schrodinger」)。在普通晶体中占据格点的原子或基因是按周期性规律发生的,例如,在普通的盐的晶体构型中,钠原子和氧原子是变化的。与此相对照,染色体是不同的原子团即基因的结晶状构型。
基本生物单位群包括病毒、基因和其他某些有繁殖能力的系统,诸如近年来常常讨论的细胞质基因,可能还有抗体。这里存在着三个基本问题。第一个是将这些系统以特定的方式结合成一体的力的问题。从物理学的观点来看,这些系统是巨大的,因为它们包含着几百万个原子。大分子处于溶液分子不断碰撞的过程中,处于热运动中。尽管如此,基因仍是非常稳定的结构。基因和染色体只要不发生遗传的变异或突变,就会在许多世代中无变化地传递下去。单个基因的突变频率几乎没有什么规律。
第二个问题是在这些单位中生长的条件问题。它们的生长不能同普通的聚合作用相比,例如不能与在合成橡胶生产中的聚合相比。因为后者分子纵向地连接起来,以致使链的长度增长,最终会横向地发生断裂。而线状的病毒分子必须横向地添加上合适的构成单位,使得最终发生纵向的分裂,正像我们在染色体分裂中直接看到的那样。可是,普通晶格力决不能充分说明基本生物单位具有的令人惊奇的特异性:它能从可用的组成物质中挑选出“恰当的成分”,把它添加到正确的位置上。这些特殊的吸引力可以直接被观察到。正如人们熟知的。动物或植物的每一个体细胞都具有两组染色体(二倍体),即每种染色体有一对;在性细胞成熟期间,染色体通过减数分裂的方式进行分配,结果每个性细胞只得到一组染色体(单倍体),以致在受精时通过卵细胞与精子细胞结合,重新组成二倍体。减数分裂的特殊阶段是染色体的配对。两条染色体互相并列地靠在一起,每对的成员通常互相缠绕,由此会发生染色体片段的交换,并因此而发生交叉。配对并不都发生在同源染色体即形态上对应的二倍体组成员之间,也发生在同源染色粒即表示基因位点的染色较深的片段之间。例如,含有染色粒的染色体的部分,可以用X射线打掉。如果这样的染色体与正常染色体配对,那么,由于同源染色粒之间发生配对,以及在一条染色体中缺失一个片段,结果由完整的染色体形成为一个环。至于由基本生物单位及其部分所施加的特殊吸引力的本质,目前只能作假设陈述。按照弗里德里希-弗雷克萨(Friedrich-Freksa)的看法,这些特殊吸引力是由核酸链的静电荷构型产生的,而约尔丹(P.Jordan)则认为,是由量子力学的共振引起的。
第三个基本问题是基本生物单位具有进行协变复制的能力。有人说过:“在这不可测知的自然奥秘面前,我们不能不感到敬畏。”(弗雷一维斯林)但是,事实上人们已用若干方式对协变复制理论进行了探讨。作行已建立了一个假说模型,可在此作一简要介绍。
德林格尔(Dehlinger)和韦茨(Wertz)(1942)已把由冯・贝塔朗菲建立的“开放系统的稳态”理论应用于基本生物单位。按照他们的看法:“符合贝塔朗菲假设的最简单的排列,即尽管处于准静止态,但不断进行化学反应的排列,是所谓单维晶体,这种排列是由不同数量的分子(原子团)一个衔接一个地构成的,它从外界吸取分子进行扩增,而且它能够进行分裂。”这个概念被详尽地阐发为更加明确的基本生物单位的模型(冯・贝塔朗菲,1944年)。按照冯・贝塔朗菲的看法,基本生物单位是微晶,一方面微晶依靠特殊的吸引力把分子团连接在一起,由此而生长,另一方面微晶又经历着分解代谢的过程。如果出现这样的过程,必定会产生排斥力,这最终会导致微晶的分裂,即导致它的协变再复制。
这个基本生物单位模型的基本假定是,它们不是稳定的晶体,而是像所有生物系统那样,处于连续的物质交换的状态中。至少,对于染色体来说这个概念基本上是必不可少的,而且在实验上得到了证实。正如去核细胞不能长久存活的事实所表明,染色体控制细胞的生理过程。作为遗传因子的载体,染色体对细胞和整个有机体产生直接的影响。由基因引起的化学作用表明,细胞和有肌体是新陈代谢的单位。实验的证据得出了相同的结论。从现代概念(卡斯珀森[Caspersson」)来看,核蛋白是细胞中蛋白质最重要的合成中心。赫维西(Hevesy)利用放射性磷对细胞的核蛋白研究的结果表明,核酸处于不断地耗损和更新的状态中、布拉赫特(Brachet)(1945年)认为,这个概念是新颖的和重要的:染色体物质看来处于不断更新的状态中,而且它是新陈代谢的基点。因此,我们关于染色体是“产生代谢变化的晶体”的假定,无疑是正确的。至于病毒的代谢情况,我们已指出了可用实验证实的方法。
从这个概念可以引出两个普遍性的结论。第一,基因和染色体看来不是静态的大分子或这些大分子的复合物,而是动态的结构,“产生代谢变化的非周期性晶体”。它们依靠一种并非静止的、以稳态方式维持的构型而存在。
第二,人们常常讨论病毒是“生命有机体”还是“无生命的自催化剂”的问题,可以用以上概念来解答。最近的研究表明,“病毒”是关于具有各种不同性质的实体的集合名称。它包括大蛋白质分子,例如烟草花叶病毒;分子束,如昆虫的多面体病毒;可在电子显微镜下看到的随着分化而形成的病毒,如疫苗病毒;最后是已接近细菌构造的形态,诸如污水微生物和引起斑点热病的立克次氏体属微生物。我们可以只限于考察像烟草花叶病蛋白质那样的形态,因为它可能代表一种产生代谢变化的微晶。这种微晶能够从它周围吸取分子团,由此而生长,最终产生分化。但它没有自主生活即进行有机分子初级合成的能力。它肯定缺乏有机分子初级合成的必要条件――酶系统。因此,基本生物单位表现出生长和协
变复制,而只有具备合成活动即构成新的有机分子的完整复合物,才能保持细胞作为一个整体的优势。
让我们来考察作为生命基本单位的细胞。细胞,即其基本组成部分是核酸和细胞质的单位,是已知的能够自主生活的最简单的系统。非常令人惊讶的事实是,一切生物,从微小的单细胞水藻到千年大树,从阿米巴到人,都由细胞这样一种构成单位的变异和多种多样的组合而成。这个事实表明存在着基本结构定律。
从物理-化学的观点来看,原生质是非常复杂的胶体系统。在这系统里,弥散在水中的状态,可以由大小极为不同的粒子、聚集状态、物理性质、化学构造和生理活动来描述;其中,弥散的程度是极易可变的,也极易从溶胶变为更密实的凝胶状态。
然而,显而易见的是,即使最复杀的胶体系统仍没有显示出“活的”东西的奇特行为,这种行为持证是,它不像普通物理-比学系统那样,尽可能快地进入稳定平衡的状态,而是在新陈代谢的稳态中保存自身(p.125)。于是,我们遇到了原生质特殊组织的问题。广泛被人们接受的现代概念,是原生质网状结构的概念(弗雷-维斯林);亚显微粒子通过它们节点或“接合点”上的侧链的连接,而形成不稳定的纤维状结构、长线似的分子、尤其是蛋白质分子(哈弗特庞克特[Haftpunkt])。哈弗特庞克特理论确实阐明了原生质组织的一个重要原理,解释了原生质的许多物理-化学性质。
可是,静态的和结构的原生质概念,看来并没有提供出一个完备的解决办法。我们必须考虑前面大体上论述过的观点(pp.16f.),注意到从结构方面进行解释的局限性。
大约四十多年前,霍夫迈斯特(Hofmeister)(1901年)试图回答细胞中活动过程的模式问题。例如,在体积大约是针头的十万分之一的肝脏细胞中进行着大量复杂的化学过程;化学家要造成这些化学过程,必须采用大量的步骤,如果我们假定化学家确实能完全完成这些过程的话。但是,细胞则几乎是靠专一的酶完成这些过程。霍夫迈斯特根据当时流行的原生质结构泡状学说,解释了细胞内不受干扰地、有序地进行的维持生命的比学反应活动:原生质由泡状结构再分为极小的单独反应的容器。这种认为小泡或泡沫状的结构在原生质中是基本的、普遍存在的观念,现已不再保留。然而,当我们考虑到在其他结构状态中发生的单独的、有序的反应过程,诸如酶依靠化学力、吸附力、电力或其他亲和力,固定在某些显微结构中或分子结构的某些点上进行反应活动,这种基本观念迄今并没有改变。
虽然,许多事例表明了酶固定在原生质显微结构和亚显微结构中,但这并没有对细胞中发生的反应模式作出最后的解释。原生质经常处于变动不居的状态,它的流动和运动已表明了这一点。它的胶质结构是非常易变的。它的体积可以通过“帽状质壁分离”的水合作用而增加十信。甚至更多;但它仍是活的,并能够逆向脱水(赫夫勒[Hofler」)。这与永久性的分子结构的存在状态几乎没有相同之处。用经过离心和分割处理的卵做实验,证明即使它处于严重分离的状态,细胞质的显微结构和亚显微结构被破坏,并不一定造成发育的紊乱。尤其是,原生质在它的物质连续不断的令成和分解中维持其自身,这个过程以结构的持续和有序的变化为先决条件。
原生质的组织结构不是静态的,而是动态的。这种动态过程固有的有序性,并不是由预先确立的结构状态造成的。相反地,作为整体的动态过程自身就具有有序性,它表现为自我调节的稳态。因此,当原生质系统尚未形成固定的结构状态时,看来它对于外界扰动有很大的忍受能力;但是,如果固定的结构状态形成了,例如,在胚胎发育过程中原生质分化成许多器官形成区域(P.58),那么,当这些状态受到不可逆的破坏或易位,就会导致系统无可修复的结果。
某些事实已表明了这种概念是正确的,尽管这样的事实目前还不多。很可能个别的组分不是以确定的化学个体存在于细胞中,而是与大量组分处于动态平衡之中。因此,泽伦森(Sorensen)认为细胞蛋白质并不严格地具有可确定的分子,而是表现为一个“组分可逆的、可分离的系统”。这个系统依赖于存在的条件,可以分解为碎片,也可以由这些碎片重新加以组合。弗莱斯(Vles)和盖克斯(Gex)用紫外线光谱仪研究透明的海胆卵。他们获得的光谱与蛋白质光谱不相一致。典型的蛋白质光谱只有在细胞被破坏之后,也就是将细胞溶解在稀释溶液中或将细胞压碎之后,才能出现。除非我们把蛋白质不看作是稳定的化合物,不看作是稳定化的产物,也不看作是不存在于活细胞中的组分,而是把它看作处于某种动态平衡中的组分,否则,上述结果很难得到解释。很有可能,细胞结构至少部分不是自发的”结构,即不是建立在稳定的物理-化学平衡基础上的结构,而是需要供给能量以维持它们存在的非自发的结构。很久以前,迈耶霍夫(Meyerhof)强调了细胞分裂时明显消耗的能量与通过胚胎呼吸所获得的能量之间的不均衡性。他得出结论认为,我们可能只是部分地知道细胞实际做的功,并且认为这种微小空间范围内所做的功对于维持细胞的结构是必要的。
因此,原生质组织是从结构基础上的有序问题进入到稳态的维持问题的一个交汇点。今后的有关理论将必须考虑到这两方面问题。
“细胞理论”是不适当的,这种陈述是“整体论”生物学最流行的一种陈述。为了对细胞理论作出正确的评判,有必要先弄清它的含意。
细胞,即由细胞质和细胞核这两个基本部分组成的系统,是所有机体(植物和动物)的最重要的结构要素。这是既无可争议而又无须称为“理论”的经验事实。这种经验陈述同有关细胞结构和功能的所有特定的经验事实一起,可称为“细胞学说”。
可是,“细胞理论”比这种经验陈述更进一步。从形态学方面说,它意味着细胞是生命世界无所不在的和唯一的构成要素,也意味着多细胞有机体是诸细胞的聚集体。从胚胎学方面说,它把多细胞有机体的发育解析为胚胎中诸个体细胞的活动。从生理学方面说,它把细胞看作功能的基本单位。细胞理论的奠基人施旺(Schwann)早在1839年就提出这样的问题;是有机体的总体决定了它的组成单位细胞的生长和发育,还是正相反,是细胞的基本力量决定了有机体?他选定了后一种看法。
先让我们来考察形态学的陈述。“所有有机体都由细胞组成”这种习惯断言如果是以绝对肯定的方式表达的,那它就是不正确的。复杂的原生动物,例如纤毛虫,长期被称为“细胞”的动物。例如,草履虫具有类似存在于高等有机体中作为多细胞系统的器官的单细胞器;细胞嘴和肛门,有收缩性的结构和神经原纤维结构,运动的器官,等等。因此,单细胞有机体的细胞只与作为整体的多细胞有机体类似,而与多细胞有机体的个体细胞不相类似。事实上,大自然已多次做了创造没有细胞分化的较大有机体的实验。SiPhoneoe,一群绿海藻便是例子。海生物种通常有几米长,具有蔓延的“茎”,细细分叉的“根”,各式各样羽状和圆齿状的“叶”,但这整个有机体是由单个巨大的多核细胞构成。确实;这样的形态是非常稀少的,这表明,在大自然中,这种非常规细胞的设计显然是经不起考验的。细胞的分化提供了重要的功能性结构,尤其造成了细胞表面得到大发展的优势。由于细胞的物质交换是在其表面进行的,所以非常规细胞组织与细胞组织相比,处于不利地位,是可以理解的。细胞分化还促进功能的分化;而另一方面,细胞膜和植物细胞的充盈,起着重要的力学作用。这些思考使我们能够理解自然界为什么顽强地表现出细胞结构向更高级的组织形态进化的天性。但是,即使较高等动物有机体也并不是无例外地由细胞构成的。非细胞形态的其他结构随处都有,例如,没有组织成典型的细胞但形成多核团块的原生质(原质团和合胞体),肌肉纤维,神经纤维和结缔组织的纤维,粉状物或胞间质,体液,等等。因此,比较高等的有机体不能简单地称为“细胞群体”。
这些类似的考虑也适用于细胞理论的其他两方面问题。多细胞有机体的发育不是诸细胞活动的总和,而是胚胎作为一个整体的活动,无论单细胞阶段还是多细胞阶段,都是如此。这种整体的胚胎活动表现为调整、定型和形态发生活动(PP.43f,57f,63)。从生理学上看,有机体的整体决定细胞的活动,而不是细胞的活动决定机体的整体。功能的分化不是由细胞决定的,而是由器官决定的,功能可以是属于细胞的部分或细胞的,也可以是属于细胞的复合体的(海登海因[Heidenhain])。
我们所思考的有机体的组织结构,是一种不仅普遍存在于生物学领域,而且也广泛存在于心理学和社会学领域的典型型式。这种型式可称为等级秩序。伍杰(Woodger)运用数理逻辑对等级秩序的原理作了规定。
可以用这样的例子来说明抽象意义的等级秩序:一个方块分成四个小方块,再把每个小方块分成四个更小的方“块,等等。这意味着,事物W处于与项或“成员”M的关系中,而这些项或“成员”与更小的项或“成员”又处于关系R中。在上面所举的例子中,R表示这样的关系:“是上一个层次成员的四分之一部分”。一个“层次”就是一类成员,它们使W处于R的同一“势力范围”之内。伍杰举出了以下的生物学实例:
Ⅰ.分化等级,即从细胞的分化及其派生细胞中产生的细胞的四维秩序。关系R(d)在这里表示为:“直接派生细胞”。W代表母细胞;细胞第一代、第二代……等等,代表等级的第一层次、第二层次……等等。
分化等级有两类:(a)这类中所有成员即细胞,是独立的有机体(原生生物);(b)这类中只有第一个成员即受精卵代表独立的有机体,而所有其他成员保持相互联系,由此形成有机整体(多细胞植物和动物)的各个部分。
Ⅱ.在Ⅰ(b)中产生多细胞有机体的空间等级,这个空间等级由各个部分的等级秩序构成,这些部分连接成逐级上升序列的系统。这里W代表整个有机体,M是有机体的组分,R(s)代表一个组分对应于下一层次某个组分的组织关系。
在所谓有机系统的“部分”中,应区分两类“部分”。一类是“组分”(component),它是各个部分的集合,相对于部分而言,组分处于关系R(s)中。因此,细胞核、细胞、组织和器官是组分,我们可以区分为:(a)构成细胞的组分,(b)细胞,(c)由细胞构成的组分。另一类是“成分”(Constituent),它是处于空间等级之外的部分,即它不能分析为更小的组分,例如软骨或骨的粉状物,结缔组织的小纤维,血浆,蛋黄,分泌颗粒,等等。成分总是“死的”。
然而,关于“成分”的定义似乎太局限了。处于细胞分化等级之外的,不一定也处于有机体空间等级之外。因此,例如结缔组织的细胞间物质不是分化等级的成员,即它处于关系R(d)之外,但它的部分可以分析为有等级层次的组分,诸如不同层次的纤维系统,小纤维,胶态分子团的组合,等等。正如海登海因的组织系统概念所表明的(PP.41f.),不仅细胞组分和细胞,而且由细胞构成的组分,都具有分化的能力。第二,细胞间物质和其他形成物并不处于组织关系R(s)之外。除了细胞以外,细胞间物质也是更高层次组织结构的必要组分。尤其在植物中,我们可以发现许多“死的”结构,诸如,细胞膜,软木,管胞和导管等等。然而这些是“活的”有机体的必要组分。化学的和无机的部分,诸如水、激素、离子,即使当它们不是细胞的组分而是处于体液中时,也必定属于有机体系统,即它们共处于关系R(s)中。尽管细胞是自主生命的最基本的单位,但多细胞有机体不独是一种细胞等级体系。
这些思考对于判断组织学中有很大争论的问题即细胞间物质的意义问题是重要的。动物有机体的支撑组织――结缔组织、软骨、骨、牙质等等――存在于大部分细胞间物质之中,而诸多细胞则嵌于细胞间物质之内。这里我们介绍两种对立的观点。第一种观点认为,应当把这些物质看作细胞的死的分泌物,而只有细胞才是活的;第二种观点认为,细胞间物质是由活的原生质变化形成的,被保留的“生命团块”(living mass)这个概念,不仅包括细胞,而且包括细胞间物质。冯・贝塔朗菲(1932年)指出,从机体论概念的观点看,第一,细胞间物质的生长和形态发生不足以断定它们的自主“生命”;第二,它们的形成不是个别细胞活动的总和,而是整体(通常是同体原生质、组织)的一元的活动;第三,生命团块的概念应考虑用系统概念来代替。在有机体的等级秩序中,首先是细胞,然后是组织,是“活的”,在组织的结构中,细胞间物质起着类似于细胞膜或小纤维在细胞中的作用,就其本身而言,它们也不是活的,但它们属于细胞系统,它们作为一个整体是活的。组织学新近的发展,尤其是赫泽拉(Huzella)关于细胞间组织的学说(1941年)证实了这种机体论观点。传统的细胞理论是片面地根据对个别细胞的结构和功能的研究建立起来的,它不能解释确定结构的统一整体与和谐的协同活动是怎样从由卵细胞分化而成的细胞聚集形态中产生的。另一方面,“极权主义的”(totalitarian)概念作为第一种观点的对立面而出现。它忽略细胞的个体性,把细胞、细胞间物质、小纤维等等,看作“生命团块”的合胞原生质连续体。与细胞理论相一致的是,细胞间组织理论强调细胞是最基本的自主生命单位,它拒绝生命团块的概念和细胞外原生质的概念。另一方面,细胞间结构是有机体整合和整体性的重要基础。无生命的细胞产物,就其本身和它们非间断的连续性而言,它们在活细胞之间的关系中起着介体的作用。按照赫泽拉的看法,小纤维和细胞膜的“弹性运动系统”,在个体发育和系统发育的早期阶段,表现为嗜银纤维(即由于硝酸银而坚韧的最细的小纤维系统),除了它具有作为支撑系统的功能之外,还起着迄今未知的建构和整合作用。它形成了细胞得以保护的生活环境;它是营养物质和液体的仓库,是细胞间关系的介体。它为细胞的有序排列,进而为形态发生提供了构架。模型实验已表明了嗜银系统的细胞外起源、无生命性质和形态发生作用。小纤维和膜结构可以从结缔组织的萃取物中产生,可以依赖细胞而生存,也可以像组织培养那样在体内生存。用适当的盐溶液与小纤维物质的溶液混合,以形成结晶,由此可以产生晶体的小纤维骨架。然后将这种纤维状物加以冲洗以消除其中的盐,用它作为培养组织生长的构架,这时它可以依靠活细胞而生存,形成最初的晶体排列。最后,细胞间的系统是生理整合的基础。例如,它能解释功能的适应性,这种适应性是由于机械张力使小纤维按一定方向形成的结果。在伤口愈合过程中,伤口充满液体,是由发炎和溶液中某些小纤维物质酸化造成的;后者产生小纤维的构架,用以作为重新长出肉芽组织的细胞的移动路线。因此,应当按照细胞间组织理论的观点修正细胞病理学。疾病的原因不能完全被归结为个体细胞的紊乱,它在很大程度上是由细胞间系统的紊乱引起的。例如,在恶性肿瘤的渗透生长过程中,细胞间系统的紊乱起着重要作用,在结缔组织中形成的小纤维大量地围住癌,促进和加剧癌细胞的侵袭。
Ⅲ.伍杰分析的等级秩序的第三种情况是遗传等级体系。这里,受精卵代表第一层次,连续几代的后代代表下几个层次。关系R(g)表示:“是直接的后裔”。可是在两性繁殖中,遗传等级体系只是更复杂的秩序系统的一个方面,因为受精卵处于双亲的关系R(g)中,该秩序系统具有网络的特征。
Ⅳ.海登海因已表述了类似的组织原理。在他看来,有机体是由组织系统构成的,这些系统按圈形上升秩序排列,上层的系统包括了下层的系统。例如,在神经中,下面的组织系统被“囊括”进另一个系统中:最低层次是神经原纤维,接着是神经元,最后是宏观神经。组织系统根据它们经分化而增殖的能力加以区分。经分化而增殖的过程不仅适用于像染色体、核、叶绿体等那些细胞组分(当然适用于细胞),而且适用于组织的细胞系统。当这样的组织系统在分化之后没有分离而保持着联系时,它们形成了越来越高等级的系统。腺单位或腺节是个例子。分化并非完全分离,它们表现出逐渐的分叉,由此最终导致树腺的形成。在许多腺类型器官(诸如绒毛、味蕾、肾脏等)中,可以发现这种“分化和综合”的原理。这个原理也适用于植物的叶,按照海登海因的说法,叶的各种各样的形状可以根据几何形结构推知。
Ⅴ.有机体不仅表现出形态学的部分等级体系,而且表现出生理学的过程等级体系。更准确地说:“一个有机体并不表现为可以从形态学方面粗略地描绘的一种等级体系。相反地,它是许多方面交织、重迭的等级体系,这个等级体系可能与形态学等级体系的层次相一致,也可能不一致。例如,有可能在动物的活动行为中发现以下的几个层次:第一,在肌肉中发生的物理-化学反应;第二,由此发生的肌肉收缩;第三,往返于脊髓中某些神经中枢的简单反射;第四,大群肌肉的复合反射,例如链反射,合作肌和对抗肌的活动,等等;第五,趋激性反应,即这样一种反射活动:躯体一侧的运动器官受到外界影响后,有机体转向针对刺激来源的一个确定位置;第六,受神经系统最高中枢的控制和统辖的整个躯体反应,它们协调所有单个活动,并且还能把这些协调活动与以前的经验联系起来;第七,依存于超个体单位的社会性反应,例如,昆虫群体中的个体活动。
过程等级体系不像形态组织那么严格。如果某个过程影响形态学方面的某种确定的组分,过程等级体系对能与形态组织相对应,但并不必然如此。某些组分,例如,胰腺的大部分组织和胰岛一起构成一种较高级的组分,构成称为“胰”的器官。但是,就其他关系而言,一个组分可以与形态学上相距很远的另一个组分协同作用,由此形成更高序列的功能系统。例如,胰岛细胞与肝脏协同作用,依靠胰岛素调节肝脏释出糖并输入血液。
这种见解是相当重要的,因为它导致这样的结论,即有机体内存在着并不构成形态学单位的“器官”。鉴于传统解剖学以形态结构为基础,现代解剖学则偏重从“功能系统”(本宁霍夫[Ben-ninghoff」)方面进行描述。行为系统,诸如由骨、肌肉和神经组成的运动系统,只有从它们的相互作用方面考虑才可理解。甚至可以说,现代解剖学最重要的进展,就在于发现这样的功能系统,例如由阿朔夫(Aschoff)和其他人发现的网状内皮系统和心脏起搏点系统的功能。
Ⅵ.等级秩序更重要的一种类型可以称为等级分异。可以在胚胎发育中看到这种最明显的例子。发育卵,最初表现为一元系统,后来逐渐分异成个别的“域”,先形成器官的复合体,随后形成个别器官、器官的部分,等等。因此,外胚层和内胚层是在作为一个整体的胚胎内形成的。在外胚层中形成了预定的表皮部位和髓板;在预定的髓板里形成了脑和脊髓部位;在脑的区域,形成了眼睛的原基,等等。用等级秩序的术语说,这里事物W对应于最初的一元的卵,下面的层次对应于第一、第二、……等等的分异序列。值得注意的是,分异并不与分化等级中的细胞组织相一致。就镶嵌卵(p.58)而言,在尚未分裂的卵内出现了分异,就调整卵而言,分异是在多细胞复合体内出现的。由于发育卵通过分异再分成细胞的组分,并没有出现决定各个部位未来结局的因素;更确切地说,这些因素是决定一组细胞变成某种组分的动力前提。例如在调整实验中,细胞的切除、移位、附加等等,并不改变器官原基的分异,则表明了这点。
分异等级体系是生物领域的主要特征,也是心理领域和社会领域的特征。物理系统的等级秩序是由最初分离的系统整合为更高级的单位而形成的,例如,在晶体中表现为空间点阵是由原子整合而成的。与此相对照,在生物领域里,由原初的整体分异为子系统。在胚胎发育过程中可以发现这种情况。系统发育过程也是这样,有机体的逐渐分化意味着最初在单细胞中联合的生命功能,分异成为摄食、消化、对刺激的反应、生殖等独立的系统。在心理学领域也有类似的情况。经典的联想心理学(PP.189f.)假定最初感受到的是对应于个别感受器要素的兴奋(例如视网膜的兴奋)的个别感觉,然后将这些个别感觉整合成知觉形式。可是,现代研究表明很可能最初感觉到的是未分异的、可以说是不定形的整体,尔后这种整体感觉才逐渐出现分异。比如,病理学的例子表明了这点。对于脑神经中枢损伤后正在恢复的病人来说,最初再现的不是单个感觉:小点光引起的感觉,不是发光点,而是一圈模糊的光亮;只有后者才是知觉到的形状,最后,逐渐恢复为发光点的知觉。这可与胚胎发育相比,视觉的恢复从未分化的状态发展到分化的状态,这可能同样适用于知觉的系统发育的进化。
Ⅶ.等级秩序的一般概念需要从各个方面加以完善。
首先,有机系统中相互作用的密切性有不同强度。在原始后生动物中,例如在腔肠动物中,细胞表现出很大的独立移动性和吞噬活动。与之相对照,我们在高等动物力发现细胞和组织对于整体严格的从属关系、我们可以称之力逐渐整合。我们进入有机体的层次越高;孤立部分的行为与它们在整体中所表现出来的行为越是不同;这些孤立部分的行为与整个有机体表现的行为相比,则更加不同。
高等动物主要有三个整合系统。第一,体液,它在组织和器官中分配营养和氧。同时为细胞的活动提供最适宜的内环境。第二,激素,它从化学上以特定的方式调节各种功能。第三,神经系统,它不仅是对环境的刺激作出反应的机构,也是有机体整合的机构。
逐渐整合与各个部分的逐渐分化协同进行,部分的分化同时意味着专一化,从形态学上说,可称为“分工”。我们可以在最简单的单细胞有机体中,也可以在最高等动物中发现新陈代谢、生长、应激性、繁殖、遗传等基本活动。可是,阿米巴的所有这些生命的基本活动过程是在单个和同一的系统即它的细胞原生质内进行,而在较高等的有机体中,这些过程分散在不同的器官和系统中进行。唯有专一化才使有机体各种功能的增强和完善成为可能;但另一方面。专一化必须为此而付出代价。
逐渐分化同时意味着逐渐机械化,即原初统一的活动分裂成诸独立的个别活动,从而失去了可调整性。当某些部分获得比较专一的功能时,失去了调整能力,即丧失了对付突然事件的其他功能。因为失去某一些部分,会造成无可修复的损坏、这个原理,可以通过社会学的类比得到最好的说明、在野蛮时代的原始公社里,每个人同时是农民、工匠、战士、猎人。只有当行业团体的成员实现专业化时,才有可能取得文化技术的进步,但这时,专业人员变得必不可缺,同时他在自己日常职业之外比原始的个人更加无能。因此,鲁滨逊在荒野中显现出的可怜相更甚于其仆人。他之所以能凑合着生存,是因为幸亏老天将各种各样的文明物冲上岸供他使用。生物领域里也是如此,发育过程中胚胎部位的逐渐确定,神经系统中随着神经中枢逐渐分化和固定,调整能力逐渐减弱,都表明了这点。同样,就有机体对不同环境适应性而言,也只有通过有机体自身内的分化和专一化,才可能得到发展。有机体必定会因为机械化而付出代价,即各个部分固定地具备单一功能,由此丧失遭受扰动后的恢复能力。
而且,随着逐渐分化,某些部分获得比其他部分更多的优势。与此相关的是逐渐集中化。于是我们在发展程度较高的等级体系中发现队列秩序原理和各个部分的从属关系(A.米勒[Muller])。整合所特别依赖的“中心”器官,在细胞中是细胞核,在高等动物中是神经系统。当然,有机体不像军队那样表现出明确的队列秩序,而是多种多样和相互作用的秩序模式的复合体。例如,我们可以把脑看作神经中枢的主导器官。但是,如果心脏停止跳动几秒钟,脑立即会变得不能活动。我们也不能认为。已脏是维持生命所必需的最重要的器官,因为当肝脏不能释出心脏活动所必需的糖时,心脏就停止跳动了。反过来,肝脏又依赖于心脏的正常活动(冯・内尔加德[von Neegard])。
队列秩序和“主导部分”的原理也是一个普遍性的原理。它不仅适用于形态组织,也适用于其他许多领域。因此,比如胚胎发育是由某些区位即组织者控制的、有机催化剂也显示出队列秩序(米塔施[Mittasch]):开始是最专一地适应进行单一反应的酶;接着是生物催化剂,诸如,植物的生长物质,或动物的组织者物质,这些物质调节不同范围的复合过程;最后是指导式的生物催化剂,诸如许多激素,它们在很大程度上影响整个有机体的心理-物理状态。同样也存在基因的队列秩序:从控制单一的、通常是微小的性状的基因,到以比较广泛的多效性(P.74)影响大量性状的基因,最后到指导许多其他基因活动的“上级”或“集合”的基因(E.费希尔[Fischer」,普福德勒[Pfaundler])。性别决定的基因属于后者;这些基因控制脊柱的遗传变化,进而不仅引起骨路系统的相关变化,而且引起肌系、神经分布等等的相关变化(屈内[Kuhne]);这些基因可能控制人的体质类型。
与集中化原理相联系的是生物个体性问题。
我们用显微镜观察一滴池塘水中的微生物,很可能会沉思这样的问题:什么是“个体”?这个问题看起来是有点多余的,实际上是深奥的和难以解答的、我们看到一滴水中活奔乱串的微小而透明的生物。绿色的像锭子形状的生物用长长的鞭毛推动自己在水中前进,拖鞋形状的生物更是自在地甩动纤毛游动着。阿米巴,无定形的原生质微滴,在泥浆中爬行。
显然,一条鱼、一只狗、一个人都是个体。我们借此意指它代表一种处于空间和时间中,具有与其他种类相区别的行为,以此经历着确定的生命循环的生物、但是,在单细胞有机体中,个体概念变得模糊不清了。单细胞生物世世代代仅以分裂的方式进行繁殖。个体意味着某种“不可分”的东西;既然单细胞生物事实上是“可分的”,而且它们恰恰是通过分裂而繁殖的,那么我们怎么可以称这些生物为个体”这同样适用于以分裂生殖和芽殖的方式进行的无性繁殖,正如我们在许多低等的后生动物中所见到的。在实验的证据面前,“个体”这个词变得不适用。当一只水螅或一只涡虫纲蠕虫可以被我们任意切割成许多片段,每个片段能生长成完整的有机体,我们能坚持认为这些动物是个体吗?用淡水珊瑚虫做实验,也证明“个体”概念是极其模糊的。将珊瑚虫的前端切开,很容易产生双头珊瑚虫。以后两个头发生竞争;如果捕获水蚤,两个头会为了战利品而发生争吵。虽然哪个头吃掉水蚤是完全无关紧要的,无论如何它总是落进共同的肠子,在那里被消化,而有机体的所有部分都由此受益。这里,我们一定要说清楚是“一个”还是“两个”个体的问题,变得毫无意义。然而,当双头动物或者分裂成两个,或者融成一个时,大自然也就回答了这个问题。
个体的概念即使在比较高等的动物中,至少在其发育的早期阶段,也成问题。不仅半个分割海胆卵可以发育成完整的动物(如杜里舒实验所表明的),而目分割的蛛螺卵也可以发育成完整的动物。另一方面,“个体”甚至可以由取自不同物种的诸部分装配而成、例如,施培曼(Spemann)将两个一半的原肠胚融合起来,产生出一个发育得很好的蝾螈,它的一侧是有条纹的,而另一侧却是有条纹和有羽饰的交杂在一起。
最后,从体质的观点来看,甚至人的个体性有时也成问题。同卵孪生儿来源于单一的卵,这个卵在胚胎的早期阶段发育成两个“个体”。人们都知道,同卵孪生儿不论其身体特征,还是其精神特征都有惊人的相似。在一对犯罪的孪生儿中,可以发现兄弟俩犯罪的性质和犯罪的时间有着惊人的一致。
因此,从自然科学的观点来看,我们只能在这样的意义上谈论个体,即系统发育和个体发育的过程中发生逐渐整合,有机体的各部分逐渐分化和失去独立性。严格地说,不存在生物学上的个体性,而只有系统发育的和个体发育的逐渐个体化,这种逐渐个体化是以逐渐集中化为基础的,某些部分取得主导的地位,由此决定着整体的行为。无论在发育过程中还是在进化过程中,个体性是可以接近但不能达到的极限。
随着个体化,死亡进入了生命世界。经验表明,高等动物中出现的复杂和整合的系统,不能以与低等生物原始的“分开”这种方式相对的分裂进行繁殖,因此它不能无限度的存在;由于自然的消耗,它们趋于衰老和死亡。用死亡的术语给个体下定义,不是不恰当的。整合系统尤其是中枢神经系统的集中化趋向,与生殖器官的分裂趋向之间,存在着直接的对抗(A.米勒)。完全的个体性,即集中化,会使繁殖成为不可能,因为繁殖以新的有机体的建构出自老的有机体的诸部分为先决条件。另一方面,恰恰是最重要的中枢系统――大脑和心脏在衰老的自然过程中最先衰弱,所以它们是死亡的器官。
因此,生物学上的个体概念只是被定义为极限的概念。确实,它来源于一种完全不同于科学和客观观察的领域。只有在作为不同于其他生物的我们自己的意识中,我们才能直接意识到个体性,但在我们周围的活机体中是无法严格定义这种个体性的。
我们见到的有机体是在空间中独特的实体。可是,它们是更高级的生命单位的成员。就时间方面说,还存在着物种的单位。每个有机体由它的同类有机体所生,并且它自身又会产生新的有机体;因此,每个有机体是超个体单位的成员。而且,就空间方面说,生命的等级体系并不终止于有机体,还存在着更高级的单位。
属于更高级生命单位的首先是同一物种的有机体的联盟,例如动物的群体。典型的例子是管水母、浮游群体的水螅虫,它们形成巨大的类族,这些类族由分化成摄食、触手、浮囊和繁殖的单位的大量珊瑚虫所构成。这种情况在动物王国中几乎是独一无二的。但是,空间中分离的有机体也可能有超个体的组织;如蚁、蜂、白蚁的昆虫社会。不同社会等级的动物,职虫、雄性与雌性、兵蚁,看来好像是昆虫社会的从属“器官”,这些“器官”很像聚合在一起的管水母群体中的各种珊瑚虫、特化动物为保持整体的生存而起协调作用的功能,正是有机体内各个细胞和器官的功能。蜜蜂在交配季节的婚飞中,在成群飞行中,在产生新皇后的过程中,整体决定了蜂窠中个体的活动,这种令人赞叹的整体的“目的性”远远超过任何个体可能的预见。整体由此得以维持和再生,例如,皇后死后,新皇后又从蜂窠中产生。因此,整体的所有标准,都适用于昆虫社会。系统发育导向组织化程度最高的动物社会的趋势,可以与导向更高等有机体的趋势相比拟;在昆虫社会中我们也发现伴随着向越来越高级和分化程度更高的组织的进步,出现了最初的松散性集合。
不仅相同有机体的联合,而且不同物种的联合都可以形成更高有序的系统。下面,我们来谈谈共生现象。共生现象也有一系列的阶段:从松散地生活在一起,如寄居蟹和海葵,到非常密切地生活在一起,如较低等的有机体通常寄生在较高等的动物特别适应的器官内。生物学证明了细胞内共生现象的广泛分布和重要性,有各种形式的共生现象,例如,营养和呼吸的共生现象,发光细菌的共生现象,等等。在某些情况下,一个新的有机体来源于两个不同有机体的共生,例如地衣是水藻和真菌的共生现象。
从由同种有机体的联合而形成的生命单位,或不同物种的共生现象,我们进到了更高级的系统。某一区域中的动物和植物群落(生物群落),诸如一个湖泊或一片森林,并不只是许多有机体的聚集体,而是受确定规律支配的单位。生物群落被定义为“在动态平衡中维持自身的种群系统”(雷斯沃伊「Reswoy」)。
最高单位是地球上整个生命界。如果一群生物体被消除,那么整个生命界必定会达到新的平衡状态或平衡被破坏的状态。只有绿色植物能够利用太阳辐射,将无机化合物合成有机物质。只有各种微生物的协同作用,保持了生物元素的循环。相似地,非常特殊的化合物,诸如对于动物的正常功能必不可少的维生素,只能由植物产生。生命之流只有在所有种群的有机体之间连续的物质流中才能维持。
正如费里德里希(Friederichs)、沃尔特里克(Woltereck)、蒂内曼(Thienemann)和其他人表明的那样,生物群落的理论属于那些整体的概念已应用了很长时期的领域。这里要谈的只是生物群落理论的一般要点。
生物群落是由相互作用的组分构成的系统,由此显示出系统的特有性质,例如彼此相互依存,自我调节,对扰动的适应,趋近于平衡态,等等。然而,生物群落整合的程度与有机体相比较,毕竟还较低些;它们是松散的、非集中化的单位。它们的发展取决于外界条件,而有机体的生活循环却是由内部条件决定的。因此,生物群落完全可称为系统,而不能像人们通常所说的那样称之为超有机体。
在生物系统中,生物群落受严格的规律控制。事实上,种群系统的数学理论,生存斗争的数学理论,生物平衡的数学理论(洛特卡[Lotka」、福尔特拉[Volterra」、德安康纳[d’Ancona」等)属于数学生物学的最先进的领域,虽然实验迄今还没有完全与这个领域的理论同步发展。在争夺食物的生存斗争中或在捕食与被捕食关系中共存的,由单一物种构成的种群的生长规律,由若干物种构成的种群的生长规律,都能由此而得到表述。
适用于生物群落的整体性概念,不仅具有理论意义,而且具有很高的实践价值。未开发的自然界处于生物群落平衡的状态。虽然,每一小片看来平静的森林或池塘中进行着生存斗争,但是,植物和动物,被捕食的动物与捕食动物之间保持着平衡;没有哪个物种能够无限地增长,因为每个物种都有自己的天敌。但是,只要不发生遗传变化或环境变化,没有一个物种会灭绝。如果人们粗心大意地干预了生物平衡,这种状态就会发生改变。他耕作土地,造成只由一种植物构成的群体,例如单调的松树林;他无意地从地球的其他地方引入了当地没有天敌的害虫。这会导致生物平衡的严重扰动。害虫如果没有天敌的控制会无限制地增长,造成完全毁灭大面积种植园的灾害。对害虫的生物控制,即用引进害虫的天敌的方法恢复生物平衡的惊人成果,表明了生物平衡理论的实际功效。
在这方面,饶有趣味的是,机体论概念适用于完全想象不到的领域即森林。莱梅尔(Lemmel)(1939年)从冯・阿塔朗菲的机体论概念中引伸出dauerwald(永存森林)观念,即避免森林净空原理和尽可能保持天然生物群落原理。
种群生长的定量定律也具有实用的价值。就人类而言,一个由单一物种构成的群体的增长,提出了社会政治的基本问题。确实,数学生物群落学是受马尔萨斯(Malthus)的人口增长学说的影响而发展起来的。流行病的循环律可以看作人、病原体和载体之间的生物平衡,它是卫生学上很重要的规律。引起动物数量周期性涨落的环境因素或种群系统自身动力学因素的问题,对于狩猎、渔业、农业和林业来说,是具有经济价值的问题。
可是,从哲学上考虑,可以提出一个深刻的问题。把一个生物群落视为统一的系统,这合理吗?难道这个系统中的成员没有卷入不断的消灭或被消灭的斗争?这导致哲学的基本问题。用罗克斯(Roux)的话来说,各个部分持续不断的斗争,在所有生物系统中――无论是在有机体中还是在生物群落系统中,都是剧烈的。不仅在“分离的”生命系统中,诸如生长中的珊瑚虫,各个组成部分为争取养料而发生竞争,而且所有生命系统都如此。例如,在饥饿的动物体内,不太重要的组织被消蚀掉,以维持更重要的组织;在再生和变态过程中,这些组织为维持整体而无畏地牺牲自己;甚至在正常发育中,造部分的分化生长,作为形态发生的一个基本过程,也是由各部分之间争夺养料而造成的。因此,由各部分竞争而形成的统一体存在于每个生物系统――有机体和超个体的生命单位中。这反映了一种可以追溯到赫拉克利特(Herachlitus)和库萨的尼古拉(Nicholas of Cusa)的深刻的形而上学观念:作为一个整体的世界及其每个个别的实体,都是一个对立的统一体,a coinci-dentia oppositorum,然而,这样的统一体在它们的对立和斗争中构成和保持了一个更大的整体。这些生物学事实展示了神正论的古老问题:世界上的罪恶产生于由个体化而成为各个竞争部分,这些竞争部分的斗争意味着个体的湮灭和整体的逐步实现。
前一页
回目录
回首页